4.
Transformation de bactéries
La transgénèse est l’addition d’un ou de plusieurs gènes étrangers dans une cellule. Le nouveau gène introduit est appelé transgène qui peut, s’il est exprimé, conférer de nouvelles caractéristiques à la cellule. La transformation de bactéries fut décrite la première fois par Griffith en 1928, puis plus tard par Avery qui démontra que l'ADN était le « principe transformant » capable de rendre des souches de Streptococcus pneumoniae virulentes.
Thèmes: Trangénèse, plasmides, bioluminescence, sélection de bactéries, résistances aux antibiotiques, ß-lactamase, formation de colonies sur boîtes de pétri.
Introduction: Dans l’expérience proposée, nous allons transformer Escherichia coli. Cette bactérie est rendue compétente (capacité à incorporer de l’ADN) par un traitement au CaCl2. Les bactéries compétentes sont d’abord gardées sur glace ce qui fige les membranes. L’ADN ajouté se fixe sur les bactéries. Un passage à 42°C restaure la fluidité des membranes permettant l’incorporation de l’ADN. Les bactéries sont ensuite cultivées une trentaine de minutes afin de permettre l’expression du gène de résistance nouvellement incorporé. Enfin ces bactéries sont étalées sur un milieu sélectif (permettant la croissance des cellules qui ont incorporé l’ADN). L’ADN utilisé est un plasmide. Il s’agit d’éléments extra-chromosomiques circulaires capables de se répliquer (de 1 à 100 copies) dans la bactérie et d’être transmis dans les cellules filles lors de divisions cellulaires. Dans la nature, les plasmides peuvent être échangés entre différentes espèces de bactéries. Ces plasmides peuvent disséminer ainsi de nouvelles caractéristiques tels que des résistances aux antibiotiques, à des métaux-lourds, ou encore de rendre les bactéries virulentes.
Le plasmide ilux: Pour cette expérience nous allons utiliser le plasmide pGEX ilux. Celui-ci contient une origine de réplication (ori), un gène de résistance à l’antibiotique ampicilline (ampR) ainsi que l’opéron lux, (un opéron est une unité de transcription) provenant de la bactérie Vibrio harveyi. Cet opéron contient 5 gènes, luxA à luxE. Ils codent pour les protéines impliquées dans la réaction de bioluminescence. Le gène ampR code pour une enzyme (la ß-lactamase) capable d’inactiver les antibiotiques de la famille des pénicillines en clivant une liaison du noyau ß-lactame. Seules les bactéries possédant le gène ampR sont capables de croître sur un milieu contenant de l’ampicilline (antibiotique de la famille des pénicillines).
Informations complémentaires concernant l'opéron ilux.

La bioluminescence: La luciférase bactérienne est une enzyme contenant 2 sous-unités (α et ß) codées respectivement par les gènes luxA et luxB. Les gènes luxC, luxD et luxE codent pour des protéines impliquées dans la conversion d’acides gras en une longue chaîne d’aldéhyde nécessaire à la réaction de luminescence.
La bioluminescence est observée dans des milliers d’espèces d’organismes vivants tels que bactéries, protozoaires, champignons, annélides, cnidaires, mollusques, insectes et poissons. Le rôle de la bioluminescence varie suivant ces organismes. On note quatre fonctions principales : 1) l’éclairage, 2) l’appât, 3) la protection et 4) la communication. Historiquement, la bioluminescence est un phénomène connu depuis l’antiquité. Aristote (348-322 av JC) parlait déjà de lumière émise par des poissons morts et des moisissures. A partir de 1600 diverses recherches permirent d’établir que cette réaction mettait en jeu une enzyme (la luciférase), un substrat oxydable (la luciférine) ainsi que l’oxygène. Les enzymes et substrats utilisés pour les réactions de bioluminescence sont extrêmement variés d’un organisme à l’autre. Chez les bactéries, la luciférase catalyse l’oxydation de la riboflavine mononucléotide réduite (FMNH2) et une longue chaîne d’aldéhyde (R-CHO), générant une flavine oxydée (FMN), un acide gras (R-COOH), de l’eau (H2O) et de la lumière (490-500 nm). La réaction est la suivante :

L'EXPERIENCE: (cliquer pour voir le schéma)
1) préparation des cellules compétentes: Le protocole présenté ici est simplifié mais permet néanmoins d’obtenir une efficacité de transformation suffisante pour l’expérience proposée. Il est important que les élèves puissent se rendre compte de toutes les étapes nécessaires au déroulement de l’expérience. Si toutefois, pour des raisons techniques, il ne vous est pas possible de préparer des cellules compétentes nous pouvons vous en fournir.
- Prendre une culture d’une nuit de bactéries Escherichia coli (DH5a). Diluer cette culture 100 fois dans 40 ml de milieu LB dans un Erlenmeyer de 250 ml. Agiter vigoureusement à 37°C .
- Après environ 3 heures, vérifier la densité optique de la culture. Quand la densité optique (OD600) est entre 0.5 et 0.6, mettre l'Erlenmeyer contenant les bactéries dans la glace.
- Prélever 1.5 ml de culture et les transférer dans 1 tube Eppendorf 1.5 ml. Marquer ce tube avec un +. Prélever 1.5 ml dans un autre tube Eppendorf. Marquer ce tube avec un -. L’ADN sera mis dans le tube marqué +. Le tube marqué - servira de contrôle.
- Centrifuger les tubes Eppendorf 30 secondes à vitesse maximale pour faire tomber les cellules.
- Jeter le surnageant.
- Ajouter 1 ml de CaCl2 0.05M froid sur le culot et le resuspendre en utilisant la pipette P1000. Attention, il faut pipeter doucement et éviter d'aspirer les cellules dans la pipette elle-même.
- Centrifuger les tubes comme avant. Jeter le surnageant et resuspendre le culot de cellules dans 100 µl de CaCl2 0.05M froid.
- Les bactéries sont gardées sur la glace et sont maintenant prêtes à l'emploi ("compétentes" pour la transformation).
- Il est important que les solutions et les tubes soient gardés sur glace.
2) la transformation:
- Ajouter 8 µl d’ADN dans le tube marqué +.
- Mettre le tube marqué + et le tube marqué - sur la glace pendant 15 minutes.
- Incuber les tubes deux minutes à 42°C (choc thermique)
- Replacer les tubes dans la glace pendant 5 minutes.
- Ajouter 1 ml de LB et incuber la suspension à 37°C pendant 30 minutes (pour l'expression de la résistance à l'ampicilline).
- Déposer et étaler:
- 0.1 ml du tube + sur une boîte LA + Amp
- 0.1 ml du tube + sur une boîte LA
- 0.1 ml du tube – sur une boîte LA+ Amp,
- 0.1 ml du tube – sur une boîte LA
- Centrifuger le reste de la suspension 30 secondes à vitesse maximale pour faire tomber les cellules.
- Enlever 0.7 ml de surnageant et le jeter.
- Resuspendre le culot avec le reste (~ 0.1 ml) du surnageant
- Déposer et étaler ces 0.1 ml sur une boite LA+Amp.
- Mettre les boîtes à 37°C pendant 24 h. Les boîtes peuvent ensuite être gardées au frigo (4°C) et incubées la nuit à 37°C juste avant de les observer en classe.
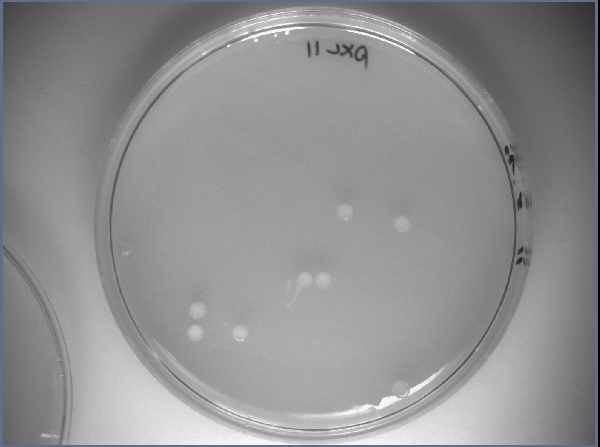
3) Résultats:
- Regarder le nombre de colonies obtenues sur chacune des boîtes et discuter des résultats
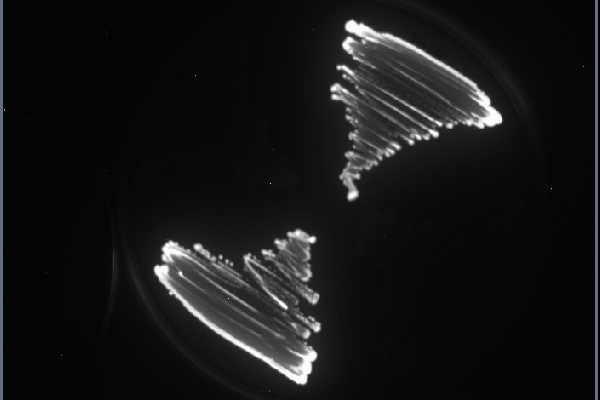
- Observer les boîtes à l’obscurité et discuter de l’expérience
Voici un exemple de ce que l'on peut observer:
|
|
|
 |
 |

Pour ceux qui désirent continuer
Comme décrit ci-dessus, les bactéries ayant intégré le plasmide non seulement resitantes à l'ampicilline mais elle sont également capable de produire de la bioluminescence. Ces deux caracteristiques nous permettent de vérifier que le plasmide a bien été intégré. Un autre moyen de le confirmer est de mettre en évidence, par PCR, un gène qui est présent uniquement sur le plasmide.
Nous allons donc amplifier ici le gène de résistance à l'ampicilline (environ 860 pb), en effectuant la PCR directement sur des colonies bactériennes.
- Préparer le mélange de réactif PCR juste avant l'emploi.
- Pour une classe de 16 par exemple, on compte 16 réactions plus un contrôle négatif ainsi qu'un contrôle positif, soit 18 réactions au total. Afin de facilité le pipetage et pour avoir assez de matériel en cas d'erreurs, nous prévoyons un mélange pour 22 réactions.
| 1 réaction | 22 réactions | |
| 2xTaq Ready Mix | 12.50 µl | 275 µl |
| Primer mix | 5.00 µl | 110 µl |
| DMSO | 1.25 µl | 27.50 µl |
| H2O | 6.25 µl | 137.50 µl |
- Préparer le mélange dans un tube Eppendorf 1,5 ml. Mélanger en pipetant en haut en bas délicatement.
- Distribuer ensuite les 25 µl du mélange dans les petits tubes PCR préalablement marqué sur le côté par les initiales des élèves.
- A l'aide d'un cure-dents, prélever une colonie isolé d'une de vos boites marquée + et la resuspendre dans le mix PCR.
- Dans le tube déstiné au contrôle positif, ajouter 1 µl du plasmidepLux117. Ne rien ajouter dans le tube destiné au contrôle négatif.
- En suivant les instructions du mode d'emploi de la machine PCR, mettre les tubes dans la machine et lancer le programme Amp..
- Le programme dure envirn 1h30 et comporte les cycles suivants:
|
94°C, 5 minutes 94°C, 30 secondes 72°C, 5 minutes |
 |
30 cycles |
- A la fin de la réaction de PCR les tubes peuvent être gardés au congélateur
- Arrêter la machine PCR en suivant les indications du mode d'emploi
- Préparer un gel à 1.5% d'agarose comme décrit dans l'expérience 10b: PCR PV92
- Récupérer vos tubes. Ils contiennent déjà le tampon de charge (rouge) et sont donc prêt à être mis sur gel.
- Charger le gel:
1er puit: 5 µl de marqueur
2ème puit et suivants: 20 µl de vos échantillons
Avant dernier puit: le contôle négatif
Dernier puit: le contrôle positif
- Allumer le transformateur et faire migrer à 100V (ampérage maximum).
- Quand le colorant rouge de migration arrive en bas du gel, arrêter le transformateur.
- Laisser refroidir le gel quelques instants. Eventuellement en le mettant au frigo. L’intensité du signal sera ainsi plus forte.
- Prendre le gel et visualiser les bandes sous la lampe bleue.
- Prendre une photo.

Cette expérience a été initialement mise en place par C. Béguin et M. Goldschmidt-Clermont. La partie concernant la PCR a été mise en place en collaboration avec Mme Starkenmann et M. Chakhparonian, enseignants au Collège Madame De Staël.
Matériel fourni:
- Micropipettes P20
- Micropipettes P200
- Micropipettes P1000
- Boîte de pointes jaunes
- Boîte de pointes bleues
- Boîte pipettes Pasteur pour les étalements
- Portoir tube eppendorf
- Mini-centrifugeuse
- Bloc chauffant
- Boite tubes Eppendorf 1,5 ml
- Plasmide lux117
- Boîtes de Petri LA
- Boîtes de Petri LA + ampicilline
- Cellules compétentes Escherichia coli
- Milieu LB liquide
- Sac jaune pour les déchets
Matériel non fourni:
- CaCl2 (en option, pour préparation de cellules compétentes)
- Matériel PCR (en option)
L’Université de Genève décline toutes responsabilités en cas de dommages survenus durant les expériences.

{kind=link}